
Вступление:
Огромная проблема современной спортивной науки заключается в том что в ней слишком много спорта и слишком мало науки. Современные профессионалы знают как научить одаренного мальчишку бегать стометровку за 10 секунд но не знают как сделать так чтобы его родители избавились от 20 -30 кг жира. Могут научить штангиста толкать и рвать за две сотни килограммов но не могут сделать так чтобы он при этом не выглядел как морж. Могут слепить очередного Дориана Ятса но не могут повторить свой успех дважды. Все теории и серьезные исследования существуют где-то далеко, в мире спорта высоких достижений, нам с вам достаются переданные через глухой телефон принципы тренировок и питания также далекие от наших с вами желаний и потребностей как и ваша машина далека от болида формулы один. Самая главная беда — отсутствие единой теории работы и развития мышц понятной, доступной и воспроизводимой на уровне любительского фитнеса.
Основа любых видов спорта это движение, а в основе любого движения лежит мышечное сокращение. Давайте для начала разберемся именно с этим феноменом. С позволения Вадима позаимствуем из его книги описание строения мышцы и мышечного волокна:
«Как известно, все ткани организма имеют клеточную структуру, не представляют исключение и мышцы. Поэтому мне придется провести краткий экскурс в цитологию — науку о клетке, и напомнить читателям о роли и свойствах основных структур клетки.
В грубом приближении клетка состоит из двух важнейших, взаимосвязанных между собой частей - цитоплазмы и ядра.
Ядро - содержит в себе молекулы ДНК, в которых заключена вся наследственная информация. ДНК — полимер, закрученный в виде двойной спирали, каждая спираль которого составлена из огромного количества четырех видов мономеров, называемых нуклеотидами. Последовательность нуклеотидов в цепочке кодирует все белки организма.
Ядро ответственно за размножение клетки - деление. Деление клетки начинается с разделения молекулы ДНК на две спирали, каждая из которых способна достроить парную из набора свободных нуклеотидов и вновь превратится в молекулу ДНК. Таким образом, количество ДНК в ядре удваивается, далее ядро делится на две части, а за ним и вся клетка.
Цитоплазма - это все, что в клетке окружает ядро. Она состоит из цитозоли (клеточной жидкости), в которую включены различные органеллы, такие как митохондрии, лизосомы, рибосомы и прочие.
Митохондрии - это энергетические станции клетки, в них с помощью различных ферментов происходит окисление углеводов и жирных кислот. Энергия, выделяющаяся при окислении веществ, идет на присоединение третьей фосфатной группы к молекуле Аденезиндифосфата (АДФ) с образованием Аденезинтрифосфата (АТФ) — универсального источника энергии для всех процессов, протекающих в клетке. Отсоединяя третью фосфатную группу и вновь превращаясь в АДФ, АТФ выделяет запасенную ранее энергию.
Ферменты или Энзимы - вещества белковой природы в сотни и тысячи раз увеличивающие скорость протекания химических реакций. Практически все жизненно важные химические процессы в организме происходят только в присутствии специфических ферментов.
Лизосомы - округлые пузырьки, содержащие около 50 ферментов. Лизосомные ферменты расщепляют поглощенный клеткой материал и собственные внутренние структуры клетки (автолизис). Лизосомы, сливаясь в фагосомы, способны переваривать целые органеллы, подлежащие дезинтеграции.
Рибосомы - органеллы, на которых происходит сборка белковой молекулы.
Клеточная мембрана - оболочка клетки, она обладает избирательной проницаемостью, то есть способностью пропускать одни вещества и задерживать другие. Задача мембраны сохранять постоянство внутренней среды клетки.
Строение мышцы.
Структурно-функциональной единицей скелетной мышцы является симпласт или мышечное волокно - огромная клетка, имеющая форму протяженного цилиндра с заостренными краями (в дальнейшем под наименованием симпласт, мышечное волокно, мышечная клетка следует понимать один и тот же объект). Длина мышечной клетки чаще всего соответствует длине целой мышцы и достигает 14 см, а диаметр равен нескольким сотым долям миллиметра. Мышечное волокно, как и любая клетка, окружено оболочкой - сарколемой. Снаружи отдельные мышечные волокна окружены рыхлой соединительной тканью, которая содержит кровеносные и лимфатические сосуды, а так же нервные волокна. Группы мышечных волокон образуют пучки, которые, в свою очередь, объединяются в целую мышцу, помещенную в плотный чехол соединительной ткани, переходящей на концах мышцы в сухожилия, крепящиеся к кости.
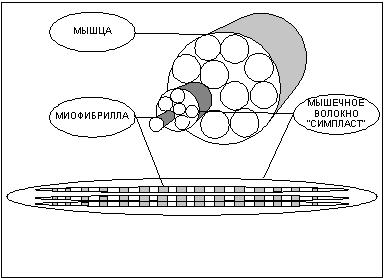
Рис.1
Усилие, вызываемое сокращением длины мышечного волокна, передается через сухожилия костям скелета и приводит их в движение.
Управление сократительной активностью мышцы осуществляется с помощью большого числа мотонейронов (рис.2) — нервных клеток, тела которых лежат в спинном мозге, а длинные ответвления - аксоны в составе двигательного нерва подходят к мышце. Войдя в мышцу, аксон разветвляется на множество веточек, каждая из которых подведена к отдельному волокну. Таким образом, один мотонейрон иннервирует целую группу волокон (так называемая нейромоторная единица), которая работает как единое целое. 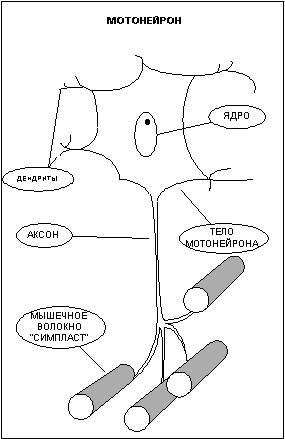
Мышца состоит из множества нервномоторных единиц и способна работать не всей своей массой, а частями, что позволяет регулировать силу и скорость сокращения.
Для понимания механизма сокращения мышцы необходимо рассмотреть внутреннее строение мышечного волокна, которое, как вы уже поняли, сильно отличается от обычной клетки. Начнем с того, что мышечное волокно многоядерно. Связано это с особенностями формирования волокна при развитии плода. Симпласты (мышечные волокна) образуются на этапе эмбрионального развития организма из клеток предшественников - миобластов. Миобласты (неоформленные мышечные клетки) интенсивно делятся, сливаются и образуют мышечные трубочки с центральным расположением ядер. Затем в мышечных трубочках начинается синтез миофибрилл (сократительных структур клетки см. ниже), и завершается формирование волокна миграцией ядер на периферию. Ядра мышечного волокна к этому времени уже теряют способность к делению, и за ними остается только функция генерации информации для синтеза белка.
Но не все миобласты идут по пути слияния, часть из них обособляется в виде клеток-сателлитов, располагающихся на поверхности мышечного волокна, а именно в сарколеме, между плазмолемой и базальной мембраной — составными частями сарколемы. Клетки-сателлиты, в отличие от мышечных волокон, не утрачивают способность к делению на протяжении всей жизни, что обеспечивает увеличение мышечной массы волокон и их обновление. Восстановление мышечных волокон при повреждении мышцы возможно благодаря клеткам-сателлитам. При гибели волокна, скрывающиеся в его оболочке, клетки-сателиты активизируются, делятся и преобразуются в миобласты. Миобласты сливаются друг с другом и образуют новые мышечные волокна, в которых затем начинается сборка миофибрилл. То есть при регенерации полностью повторяются события эмбрионального (внутриутробного) развития мышцы.
Помимо многоядерности отличительной чертой мышечного волокна является наличие в цитоплазме (в мышечном волокне ее принято называть саркоплазмой) тонких волоконец — миофибрилл (рис.1), расположенных вдоль клетки и уложенных параллельно друг другу. Число миофибрилл в волокне достигает двух тысяч. Миофибриллы являются сократительными элементами клетки и обладают способностью уменьшать свою длину при поступлении нервного импульса, стягивая тем самым мышечное волокно. Под микроскопом видно, что миофибрилла имеет поперечную исчерченность — чередующиеся темные и светлые полосы. При сокращении миофибриллы светлые участки уменьшают свою длину и при полном сокращении исчезают вовсе. Для объяснения механизма сокращения миофибриллы около пятидесяти лет назад Хью Хаксли была разработана модель скользящих нитей, затем она нашла подтверждение в экспериментах и сейчас является общепринятой.
Механизм сокращения волокна.
Чередование светлых и темных полос в миофибрильной нити определяется упорядоченным расположением по длине миофибриллы толстых нитей белка миозина и тонких нитей белка актина; толстые нити содержатся только в темных участках (А-диск) (рис.3), светлые участки (I-диск) не содержат толстых нитей, в середине I-диска находится Z-линия к ней крепятся тонкие нити актина. Участок миофибриллы состоящий из А-диска (темной полосы) и двух половинок I-дисков (светлых полос) называют саркомером. Сокращение длины саркомера происходит путем втягивания тонких нитей актина между толстыми нитями миозина. Скольжение нитей актина вдоль нитей миозина происходит благодаря наличию у нитей миозина боковых ответвлений, называемых мостиками. Головка миозинового мостика сцепляется с актином и изменяет угол наклона к оси нити, тем самым как бы продвигая нить миозина и актина относительно друг друга, затем отцепляется, сцепляется вновь и вновь совершает движение. Перемещение миозиновых мостиков можно сравнить с гребками весел на галерах. Как перемещение галеры в воде происходит благодаря движению весел, так и скольжение нитей происходит благодаря гребковым движениям мостиков, существенное отличие состоит лишь в том, что движение мостиков асинхронно.
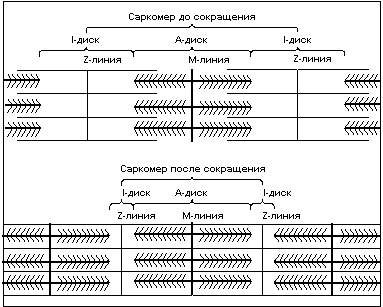
Рис.3
Тонкая нить представляет из себя две спирально скрученные нити белка актина. В канавках спиральной цепочки залегает двойная цепочка другого белка — тропомиозина. В расслабленном состоянии мостики миозина не имеют возможности связаться с актином, так как места сцепления блокированы тропомиозином. При поступлении по аксону двигательного мотонейрона нервного импульса клеточная мембрана меняет полярность заряда, и из специальных терминальных цистерн, расположенных вокруг каждой миофибриллы вдоль всей ее длины, в саркоплазму выбрасываются ионы кальция (Са++) (рис.4).
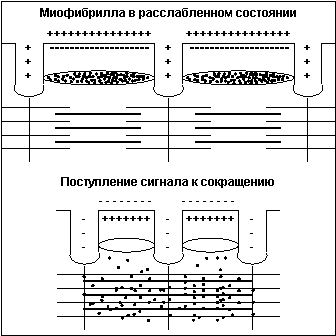
Рис.4
Под воздействием Са++ нить тропомиозина входит глубже в канавку и освобождает места для сцепления миозина с актином, мостики начинают цикл гребков. Сразу после высвобождения Са++ из терминальных цистерн он начинает закачиваться обратно, концентрация Са++ в саркоплазме падает, тропомиозин выдвигается из канавки и блокирует места сцепления мостиков — волокно расслабляется. Новый импульс опять выбрасывает Са++ в саркоплазму и все повторяется. При достаточной частоте импульсации (не менее 20 Гц) отдельные сокращения почти полностью сливаются, то есть достигается состояние устойчивого сокращения, называемое тетаническим сокращением или гладким тетанусом.
Энергетика мышцы.
Естественно, что для движения мостика требуется энергия. Как я уже упоминал ранее, универсальным источником энергии в живом организме является молекула АТФ. Под действием фермента АТФазы АТФ гидролизуется, отсоединяя фосфатную группу в виде ортофосфорной кислоты (Н3РО4), и превращается в АДФ, при этом высвобождается энергия.
АТФ + H2O = АДФ + H3PO4 + энергия.
Головка миозинового мостика при контакте с актином обладает АТФазной активностью и соответственно возможностью расщеплять АТФ и получать энергию, необходимую для движения.
Запас молекул АТФ в мышце ограничен, поэтому расход энергии при работе мышцы требует постоянного его восполнения. Мышца имеет три источника воспроизводства энергии: расщепление креатинфосфата; гликолиз; окисление органических веществ в митохондриях.
Креатинфосфат обладает способностью отсоединять фосфатную группу и превращаться в креатин, присоединяя фосфатную группу к АДФ, которая превращается в АТФ.
АДФ + креатинфосфат = АТФ + креатин.
Эта реакция получила название — реакции Ломана. Запасы креатинфосфата в волокне не велики, поэтому он используется в качестве источника энергии только на начальном этапе работы мышцы, до момента активизации других более мощных источников — гликолиза и кислородного окисления. По окончании работы мышцы реакция Ломана идет в обратном направлении, и запасы креатинфосфата в течение нескольких минут восстанавливаются.
Гликолиз — процесс распада одной молекулы глюкозы (C6H12O6) на две молекулы молочной кислоты (C3H6O3) с выделением энергии, достаточной для «зарядки» двух молекул АТФ, протекает в саркоплазме под воздействием 10 специальных ферментов.
C6H12O6 + 2H3PO4 + 2АДФ = 2C3H6O3 + 2АТФ + 2H2O.
Гликолиз протекает без потребления кислорода (такие процессы называются анаэробными) и способен быстро восстанавливать запасы АТФ в мышце.
Окисление протекает в митохондриях под воздействием специальных ферментов и требует затрат кислорода, а соответственно и времени на его доставку. Такие процессы называются аэробными. Окисление происходит в несколько этапов, сначала идет гликолиз (см. выше), но образовавшиеся в ходе промежуточного этапа этой реакции две молекулы пирувата не преобразуются в молекулы молочной кислоты, а проникают в митохондрии, где окисляются в цикле Кребса до углекислого газа СО2 и воды Н2О и дают энергию для производства еще 36 молекул АТФ. Суммарное уравнение реакции окисления глюкозы выглядит так:
C6H12O6 + 6O2 + 38АДФ + 38H3PO4 = 6CO2 + 44H(2)О + 38АТФ.
Итого распад глюкозы по аэробному пути дает энергию для восстановления 38 молекул АТФ. То есть окисление в 19 раз эффективнее гликолиза.
Типы мышечных волокон.
Скелетные мышцы и образующие их мышечные волокна различаются по множеству параметров: скорости сокращения, утомляемости, диаметру, цвету и т.д. Традиционно выделяют красные и белые, медленные и быстрые, гликолитические и окислительные волокна.
Скорость сокращения мышечного волокна определяются типом миозина. Изоформа миозина, обеспечивающая высокую скорость сокращения, — быстрый миозин характеризуется высокой активностью АТФазы, а соответственно и скоростью расхода АТФ. Изоформа миозина с меньшей скоростью сокращения — медленный миозин, характеризуется меньшей активностью АТФазы. Волокна, с высокой активностью АТФазы и скоростью расхода АТФ, принято называть быстрыми волокнами, волокна, характеризующиеся низкой активностью АТФазы и меньшей скоростью расхода АТФ, — медленными волокнами.
Для восполнения затрат энергией мышечные волокна используют окислительный либо гликолитический путь образования АТФ.
Окислительные, или красные, мышечные волокна небольшого диаметра окружены массой капилляров и содержат много белка миоглобина (именно наличие этого белка придает волокнам красный цвет). Многочисленные митохондрии красных волокон имеют высокий уровень активности окислительных ферментов. Мощная сеть капилляров необходима для доставки с кровью большого количества кислорода, а миоглобин используется для транспортировки кислорода внутри волокна от поверхности к митохондриям. Энергию красные волокна получают путем окисления в митохондриях углеводов и жирных кислот.
Гликолитические, или белые, мышечные волокна имеют больший диаметр, в их саркоплазме содержится значительное количество гранул гликогена, митохондрии не многочисленны, активность окислительных ферментов значительно уступает активности гликолитических. Гликоген, его еще принято называть «животным крахмалом», — сложный полисахорид с высокой молекулярной массой служит резервным питательным веществом белого волокна. Гликоген распадается до глюкозы, которая, служит топливом при гликолизе.
Быстрые волокна, обладающие высокой активностью АТФазы и соответственно скоростью расхода энергии, требуют высокой скорости воспроизводства АТФ, обеспечить которую может только гликолиз, так как, в отличие от окисления, он протекает непосредственно в саркоплазме и не требует времени на доставку кислорода к митохондриям и доставку энергии от них к миофибриллам. Поэтому быстрые волокна предпочитают гликолитический путь воспроизводства АТФ и соответственно относятся к белым волокнам. За высокую скорость получения энергии белые волокна платят быстрой утомляемостью, так как гликолиз, как видно из уравнения реакции ведет к образованию молочной кислоты, накопление которой повышает кислотность среды и вызывает усталость мышцы и в конечном итоге останавливает ее работу.
Медленные волокна, характеризующиеся низкой активностью АТФазы, не требуют столь быстрого восполнения запасов АТФ и для обеспечения потребности в энергии используют путь окисления, то есть относятся к красным волокнам. Благодаря этому медленные волокна являются низко утомляемыми и способны поддерживать относительно небольшое, но длительное напряжение.
Существует и промежуточный тип волокон с высокой АТФазной активностью, и окислительно-гликолитическим путем воспроизводства АТФ.
Тип мышечного волокна зависит от мотонейрона его иннервирующего. Все волокна одного мотонейрона принадлежат к одному типу. Интересный факт — при подключении к быстрому волокну аксона медленного мотонейрона и наоборот, волокно перерождается, меняя свой тип. До недавнего времени существовало две точки зрения на зависимость типа волокна от типа мотонейрона, одни исследователи полагали, что свойства волокна определяются частотой импульсации мотонейрона, другие, что эффект влияния на тип волокна определяется гормоноподобными веществами поступающими из нерва (эти вещества до настоящего времени не выделены). Исследования последних лет показывают, что обе точки зрения имеют право на существование, воздействие мотонейрона, на волокно осуществляется обоими способами.
Регуляция силы и скорости мышечного сокращения.
Как вы знаете на собственном опыте, человек обладает возможностью произвольно регулировать силу и скорость мышечного сокращения. Реализуется эта возможность несколькими способами. С одним из них вы уже знакомы — это управление частотой нервной импульсации. Подавая волокну одиночные команды на сокращение, можно добиться в нем легкого напряжения. Так, например, поддерживающие позу мышцы слегка напряжены, даже когда человек отдыхает. Повышая частоту импульсов можно увеличивать силу сокращения до максимально возможной для данного волокна в данных условиях работы, которая достигается при слиянии отдельных импульсов в тетанус.
Сила, развиваемая волокном в состоянии тетануса, не всегда одинакова и зависит от характера и скорости движения. При статическом напряжении (когда длина волокна остается постоянной) сила, развиваемая волокном, больше чем при сокращении волокна, и чем быстрее сокращается волокно, тем меньшую силу оно способно развить. Максимальную же силу волокно развивает в условиях негативного движения, то есть при удлинении волокна.
При отсутствии внешней нагрузки волокно сокращается с максимальной скоростью. При увеличении нагрузки скорость сокращения волокна уменьшается и по достижении определенного уровня нагрузки падает до нуля, при дальнейшем увеличении нагрузки волокно удлиняется.
Причину в различии силы волокна при различных направлениях движения легко понять, рассмотрев уже приведенный ранее пример с гребцами и веслами. Дело в том, что после завершения «гребка» миозиновый мостик некоторое время находится в состоянии сцепления с нитью актина, представьте, что весло после гребка так же не сразу извлекается из воды, а находится погруженным еще некоторое время. В случае, когда гребцы плывут вперед (позитивное движение), весла, остающиеся погруженными в воду после завершения гребка, тормозят движение и мешают плыть, в то же время, если лодка буксируется назад, а гребцы сопротивляются этому движению, то погруженные весла так же мешают движению, и буксиру приходится прилагать большие усилия. То есть, при сокращении волокна, сцепленные мостики мешают движению и ослабляют силу волокна, при негативном движении — удлинении мышцы — не отцепленные мостики так же мешают движению, но в этом случае они как бы поддерживают опускающийся вес, что и позволяет волокну развивать большую силу. Легче всего понять различия между статическим напряжением, позитивным и негативным движением взглянув на Рис.5.
Итак, мы рассмотрели основные факторы, влияющие на силу и скорость сокращения отдельного волокна. Сила же сокращения целой мышцы зависит от количества волокон в данный момент времени вовлеченных в работу.
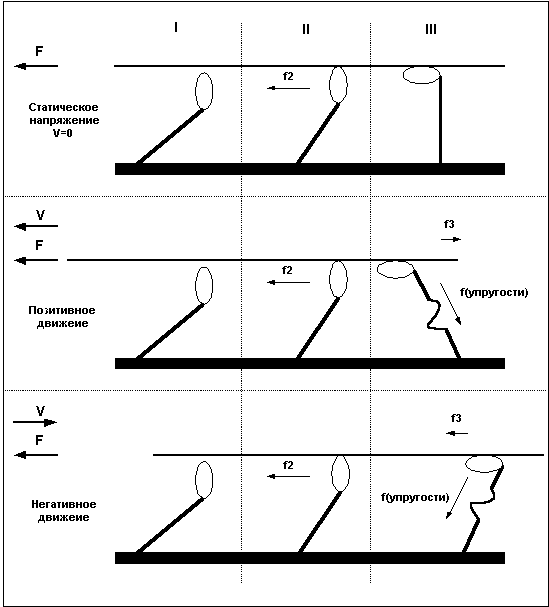
Рис.5
Вовлечение волокон в работу.
При поступлении от ЦНС (центральной нервной системы) к мотонейронам (расположенным в спинном мозге), возбуждающего сигнала (запускающий импульс), мембрана мотонейрона поляризуется и он генерирует серию импульсов, направляемых по аксону к волокнам. Чем сильнее воздействие на мотонейрон (поляризация мембраны), тем выше частота генерируемого им импульса — от небольшой стартовой частоты (4-5 Гц), до максимально возможной, для данного мотонейрона, частоты (50 Гц и более). Быстрые мотонейроны способны генерировать гораздо более высокочастотный импульс, чем медленные, поэтому сила сокращения быстрых волокон гораздо больше подвержена частотной регуляции, чем сила медленных.
В то же время имеется и обратная связь с мышцей, от которой поступают тормозящие сигналы, уменьшающие поляризацию мембраны мотонейрона и снижающие его ответ.
Каждый мотонейрон имеет свой порог возбудимости. Если сумма возбуждающих и тормозящих сигналов, превышает этот порог и на мембране достигается необходимый уровень поляризации, то мотонейрон вовлекается в работу. Медленные мотонейроны имеют, как правило, низкий порог возбудимости, а быстрые высокий. Мотонейроны же целой мышцы имеют широкий спектр значений этого параметра. Таким образом, при повышении силы сигнала ЦНС, активируется все большее число мотонейронов, а мотонейроны с низким порогом возбудимости увеличивают частоту генерируемого импульса.
Когда требуется легкое усилие, например, при ходьбе или беге трусцой, активируется небольшое число медленных мотонейронов и соответствующее число медленных волокон, ввиду высокой выносливости этих волокон такая работа может поддерживаться очень долго. По мере увеличения нагрузки ЦНС приходится посылать все более сильный сигнал, и большее число мотонейронов (а, следовательно, волокон), вовлекается в работу, а те, что уже работали, увеличивают силу сокращения, по причине увеличения частоты импульсации, поступающей от мотонейронов. По мере увеличения нагрузки в работу включаются быстрые окислительные волокна, а по достижения определенного уровня нагрузки (20%-25% от максимума), например, во время подъема в гору или финального спурта, силы окислительных волокон становится недостаточно, и посылаемый ЦНС сигнал включает в работу быстрые — гликолитические волокна. Быстрые волокна значительно повышают силу сокращения мышцы, но, в свою очередь, быстро утомляются, и в работу вовлекается все большее их количество. Если уровень внешней нагрузки не уменьшается, работу в скором времени приходится останавливать из за усталости, в результате накопления молочной кислоты.
Скорость движения мышцы зависит от соотношения внешней нагрузки и количества вовлеченных в движение волокон. Мобилизация относительно небольшого количества волокон, приведет к медленному движению, как только совместная сила сокращающихся волокон превысит уровень внешней силы. Мобилизация все большего количества волокон при той же внешней нагрузке приведет к увеличению скорости сокращения, а увеличение внешней нагрузки, или падение силы волокон в результате усталости, при неизменном количестве мобилизованных волокон, приведет к падению скорости сокращения.
При предельных нагрузках, например, при подъеме максимального веса или подъеме относительно небольшого веса, но с максимальной скоростью, сокращается сразу максимально возможное для данного индивида число волокон.
По вопросу вовлечения волокон при предельных нагрузках в литературе приводятся противоречивые сведения. Так ряд авторов утверждает, что даже у тренированных спортсменов при предельных нагрузках активно не более 85% от общего количества двигательных единиц в мышце. С другой стороны, Гурфинкель и Левин приводят данные, что большая часть двигательных единиц рекрутируется уже при нагрузках до 50% от максимума, и в дальнейшем, при повышении силы, подключается лишь небольшая часть (около 10%) самых крупных двигательных единиц, а рост силы от 75% до 100% происходит уже не за счет вовлечения новых двигательных единиц, а за счет повышения частоты импульса, генерируемого мотонейронами. Независимо от того, чья точка зрения является правильной, важно одно — сила мышцы зависит от силы импульса ЦНС, а чем обусловлен рост силы при нагрузках свыше 75% от максимальной только ли ростом частоты, генерируемой мотонейронами, или еще и вовлечением в работу новых мотонейронов не принципиально. Важно так же и то, что в мышцу встроен ограничительный механизм контроля за развиваемым напряжением, осуществляется через сухожильные органы Гольджи. Сухожильные рецепторы регистрируют напряжение и при превышении критического значения оказывают тормозящее воздействие на мотонейроны. Считается, что при отключении контроля за напряжением проявляется «сверхсила» человека, регистрируемая в экстремальных ситуациях.»
Основаы классической теорией тренировки.
Организм — саморегулируемая система, стремящаяся к поддержанию постоянства внутренней среды. Физическая нагрузка оказывает выраженное воздействие на внутреннюю среду мышц и организма в целом, смещая многие биохимические константы от уровня, характерного для гомеостаза покоя, к состоянию, соответствующему гомеостазу деятельности. Степень этих изменений зависит от характера и интенсивности физической нагрузки и индивидуальной реакции на нее организма, отражающей уровень тренированности. Сразу после прекращения нагрузки, в организме начинаются процессы, стремящиеся восстановить исходное состояние, соответствующее гомеостазу покоя, в ходе этих процессов закрепляются изменения, позволяющие в дальнейшем минимизировать возмущение внутренней среды при аналогичных нагрузках.
Спортивную тренировку можно рассматривать как процесс направленного приспособления организма (адаптации) к воздействию тренировочных нагрузок.
Различают срочную и долговременную адаптацию.
Срочная адаптация — это ответ организма на однократное воздействие тренировочной нагрузки, выраженный в «аварийном» приспособлении к изменившемуся состоянию внутренней среды и сводящийся, преимущественно, к изменениям в энергетическом обмене и к активации высших нервных центров, ответственных за регуляцию энергетического обмена.
Долговременная адаптация развивается постепенно на основе многократной реализации срочной адаптации путем суммирования следов повторяющихся нагрузок.
В развитии процессов адаптации различают специфическую компоненту и общую адаптационную реакцию.
Процессы специфической адаптация затрагивают внутриклеточный энергетический и пластический обмен и связанные с ним функции вегетативного обслуживания, которые специфически реагируют на данный вид воздействия пропорционально его силе.
Общая адаптационная реакция развивается в ответ на самые различные раздражители не зависимо от их природы, в случае, если сила этих раздражителей превышает определенный пороговый уровень. Реализуется общая адаптационная реакция благодаря возбуждению симпато-адреалиновой и гипофизарно-адренокортикальной систем, в результате активации которых в крови и тканях повышается содержание катехоламинов и глюкокортикоидов, что способствует мобилизации энергетических и пластических резервов организма. Такая неспецифическая реакция на раздражение получила название «синдром стресса», а раздражители, вызывающие эту реакцию, обозначают как стресс-факторы. Общий адаптационный синдром сам по себе не является основой адаптации к тренировочным нагрузкам, а лишь призван на системном уровне обеспечивать протекание специфических адаптационных реакций, которые и реализуют приспособление организма к конкретным видам нагрузки.
Не смотря на различную природу процессов специфической адаптации можно выделить общие закономерности их протекания. Основу специфической адаптации составляют процессы восстановления сниженных во время мышечной работы энергетических ресурсов, разрушенных структур клеток, смещенного водно-электролитического баланса и др. Наглядно проследить закономерности протекания восстановительных процессов можно на примере восстановления энергетических ресурсов организма, так как, при физических нагрузках наиболее выраженные изменения обнаруживаются именно в сфере энергетического обмена.
Мышечная работа, в зависимости от интенсивности и длительности, приводит к снижению уровня креатинфосфата в мышцах, истощению запасов внутримышечного гликогена и гликогена печени, резервов жиров. Интенсивно протекающие после прекращения нагрузки процессы восстановления приводят к тому, что в о пределенный момент отдыха после работы уровень энергетических веществ превышает исходный дорабочий уровень. Это явление получило название суперкомпенсация, или сверхвосстановление (рис. 1).
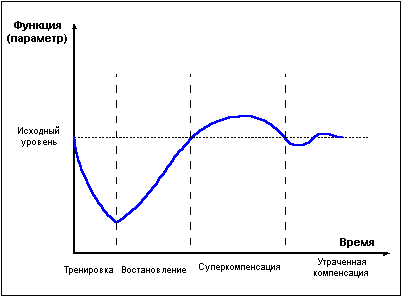
Рис.1
Фаза суперкомпенсации длится не вечно, постепенно уровень энергетических веществ возвращается к норме, испытывая некоторые колебания возле состояния равновесия. Чем больше расход энергии при работе, тем интенсивнее идет восстановление и тем значительнее превышение исходного уровня в фазе суперкомпенсации. Однако это правило применимо лишь в ограниченных пределах. При истощающих нагрузках, приводящих к большому накоплению продуктов распада, скорость восстановительных процессов замедляется, фаза суперкомпенсации откладывается во времени и оказывается выраженной в меньшей степени.
Похожим образом развивается восстановление не только энергетических, но и пластических ресурсов организма, и даже целых тренируемых функций. Напряжение в ходе физической нагрузки систем, ответственных за реализацию той или иной функции, приводит к снижению функциональных возможностей организма, затем во время отдыха достигается состояние суперкомпенсации тренируемой функции, длящееся определенное ограниченное время, далее, при отсутствии повторных нагрузок, уровень тренируемой функции вновь снижается, — наступает фаза утраченной суперкомпенсации (рис 1).
Развитие долговременной адаптации становится возможным только в случае, если достигаемый срочный и отставленный тренировочный эффект от каждой тренировки будет суммироваться по определенным правилам.
Проведение повторных тренировок в фазе утраченной суперкомпенсации (слишком редкие тренировки) (Рис. 2) не сможет привести к закреплению тренировочного эффекта, так как каждая последующая тренировка проводится после возвращения функциональных возможностей организма к исходному уровню.
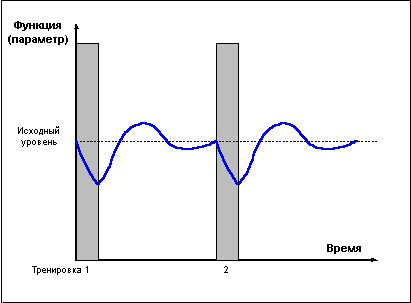
Рис. 2.
Слишком частые тренировки, прерывающие стадию восстановления до достижения эффекта суперкомпенсации (Рис. 3) приводят к отрицательному взаимодействию тренировочных эффектов и снижению функциональных возможностей организма.
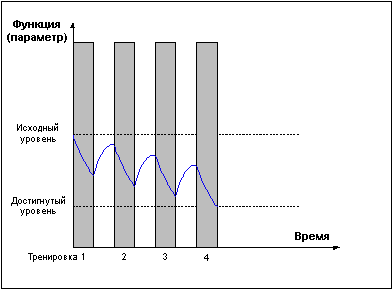
Рис. 3.
И только проведение повторных тренировок в фазе суперкомпенсации (Рис. 4) приводит к положительному взаимодействию тренировочных эффектов, закреплению следов срочной адаптации, росту тренируемой функции и формированию долговременной адаптации.
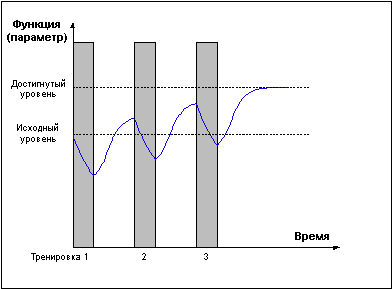
Рис. 4.
Однако не следует воспринимать приведенные выше правила слишком конкретно. Требование задавать нагрузку только в стадии суперкомпенсации справедливо лишь в долгосрочной перспективе. В рамках одного тренировочного микроцикла возможны серии тренировок в стадии недовосстановления (Рис. 5), приводящие к более глубокому истощению тренируемой функции, что может быть использовано для получения более мощного роста функциональных возможностей в стадии суперкомпенсации, либо для вызванного тактической необходимостью переноса во времени эффекта суперкомпенсаци.
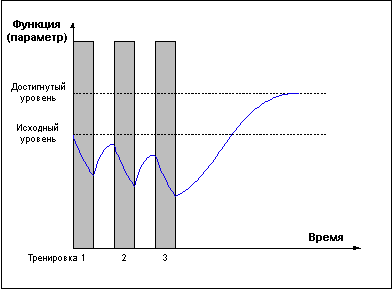
Рис. 5.
На первый взгляд может показаться, что составление эффективных тренировочных программ является делом не сложным. Достаточно определить уровень нагрузки, необходимый для достижения максимальной суперкомпенсации той или иной функции, и время наступления фазы суперкомпенсации, а далее задавать повторные нагрузки с необходимой частотой, постоянно получая положительное суммирование тренировочных эффектов. На самом деле построить тренировку по такому принципу невозможно. Дело в том, что различные параметры и функции, вносящие свой вклад в общую тренированность, необходимую для того или иного вида спорта, имеют разное время восстановления и достижения суперкомпенсации и разную длительность фазы суперкомпенсации. Так фаза суперкомпенсации креатинфосфата достигается через несколько минут отдыха после нагрузки, приводящей к существенному снижению его уровня. Для достижения выраженной суперкомпенсации содержания гликогена в мышцах требуется не менее 2-3 суток, к этому моменту уровень креатинфосфата уже вступит в фазу утраченной суперкомпнесации. А вот для восстановления структур клеток, разрушенных в ходе тренировок, может потребоваться еще больший период времени, в течение которого уровень гликогена в мышцах уже может вернуться к исходному уровню. В этой связи хочу сразу обратить внимание читателей на тот факт, что заявления многих «гуру» бодибилдинга о том, что время восстановления мышцы после тренировки должно составлять n часов/дней, без указания о восстановлении какой ведущей функции идет речь, кажутся довольно сомнительными. Задать определенный период отдыха между тренировками, позволяющий получать развитие всех тренируемых функций одновременно, невозможно.
Поэтому в классической спортивной школе годичный (и даже многолетний) период тренировок разбивают на микро и макроциклы в ходе которых ставятся задачи по развитию определенных тренируемых качеств, чередование тренировочных занятий в ходе микроциклов осуществляется таким образом, что бы физические нагрузки, направленные на развитие определенного двигательного качества задавались через промежутки времени, обеспечивающие суперкомпенсацию ведущей функции, а нагрузки иной направленности, применяемые в этот период, не оказывали отрицательного влияния на восстановление основной функции. Однако такой метод срабатывает только при развитии взаимонезависимых функций или параметров. В случае если определенное двигательное качество в равной мере зависит от развития нескольких функций или параметров, испытывающих одновременное напряжение в ходе одного тренировочного занятия и имеющих разное время восстановления, в течение микроцикла приходиться варьировать интенсивностью и объемом тренировок, накладывая волны восстановления различных параметров друг на друга таким образом, что бы получить суперкомпенсацию основных тренируемых функций к моменту завершения микроцикла.
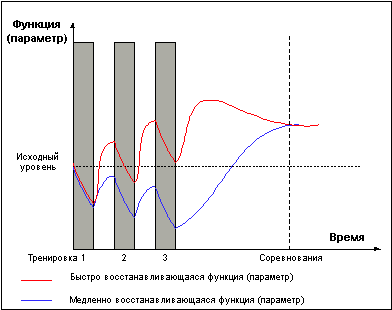
Рис.6.
На рисунке 6 представлен простейший вариант построения микроцикла для двух тренируемых функций, имеющих разное время восстановления. В течении микроцикла одна из функций испытывает последовательное положительное суммирование тренировочных эффектов, в то время как другая последовательно вводится в стадию истощения и достигает суперкомпенсации только во время отдыха, либо снижения нагрузки к концу микроцикла. Реальная картина тренировок еще гораздо сложнее, представьте, что тренируемых параметров и функций не две, а десятки. Но общие правила и закономерности, я думаю, вам уже стали понятны.
Как вы видите, классическая теория тренировки оперирует такими понятиями как двигательные качества, функциональные возможности, и основывается на изучение процессов, приводящих к росту работоспособности мышц и организма в целом в различных режимах работы. Основой практически всех видов спорта является именно работоспособность, и цель планируемых адаптационных изменений в организме спортсмена, как правило, — выход на новый уровень работоспособности. Гипертрофия мышц вовсе не является целью тренинга в классическом спорте, и воспринимается лишь как побочный продукт развития основных двигательных качеств, более того, в некоторых случаях, гипертрофия мышц может оказывать даже отрицательное влияние на достижение стоящих перед спортсменом целей. Посетителей же тренажерных залов, за редким исключением, в большей степени интересует именно достижение гипертрофии мышц, нежели развитие двигательных качеств и повышения работоспособности. Безусловно, развитие работоспособности мышц способствует гипертрофии мышечной ткани.
Процессы, направленные на улучшение доставки кислорода к мышцам существенно развивают капилярную сеть, что способствует общей гипертрофии мышц.
Тренировка окислительной активности мышц приводит к значительному росту в саркоплазме мышечных волокон количества и объема митохондрий — энергетических станций клетки.
Последовательные процессы суперкомпенсации внутримышечных запасов гликогена приводят к значительному его накоплению, что, в свою очередь, увеличивает объем саркоплазмы мышечного волокна.
Накопление иных веществ, ответственных за энергообеспечение, мышечной деятельности, таких, например, как креатинфосфат, так же увеличивает объем саркоплазмы, и даже не столько за счет объемов самих этих веществ, сколько за счет сопутствующего увеличения объема внутриклеточной жидкости.
То есть, тренировка работоспособности мышц приводит к гипертрофии мышечных волокон в основном за счет увеличения объема саркоплазмы.
Но самый существенный вклад в рост объемов и силы сокращения волокна вносит гипертрофия миофибрилл, все остальные компоненты клетки призваны лишь обеспечивать их сократительную активность. Как размер топливных баков самолета зависит от мощности его турбин, так и объем саркоплазмы мышечного волокна зависит от развития сократительного аппарата клетки (ну и конечно от объема работы, регулярно выполняемой мышцами).
Вернемся к рассмотрению процессов происходящих в мышце во время восстановления после тренировки. По завершении катаболического этапа саморазрушения поврежденных структур начинается этап компенсации — восстановления внутренней структуры волокон, который, ради справедливости должен заметить, не всегда может завершиться суперкомпенсацией. При слишком обширных травмах или отсутствии условий для восстановления результат может быть прямо противоположным.
Против теории разрушения чаще всего приводят следующие аргументы: «Если причиной роста являются микротравмы, то почему же мышца не растет, если ее бить палками?»
Ответ на этот вопрос можно найти в работе Володиной А.В., целью ее докторской диссертации является изучение процессов, препятствующих реализации регенерационного потенциала, заложенного в мышечном волокне. Эксперименты показали, что, в условиях обширного повреждения волокон, сопровождающегося ишемией (нарушением кровоснабжения) поврежденных тканей, вызывающей дефицит в снабжении волокна кислородом и питательными веществами, часть клеток-сателлит гибнет и поглощается фагоцитами, а часть идет по пути превращения не в мышечные клетки, а в фибробласты (клетки, производящие коллаген). В итоге место повреждения затягивается соединительной тканью, а количество волокон в мышечной ткани снижается, по причине гибели части из них от повреждений.
Очевидно, что при микротравмах волокон — разрушении внутренней структуры волокна без нарушения его целостности, в отличие от травм целой мышцы, снабжение волокон кислородом, а так же его иннервация не нарушены, поэтому условия, приводящие к гибели целых волокон и клеток-сателлитов, отсутствуют.
И так, если объем микротравм, полученных в ходе тренировки, был не слишком велик для срыва восстановительных процессов, но достаточен для активации клеток-сателлитов, то в подвергшемся тренировочной нагрузке волокне увеличивается количество клеточных ядер. Восстановление энергетических ресурсов после тренировки приводит к суперкомпенсации энергетических веществ, а лизис разрушенных тренировкой белков увеличивает содержание свободных аминокислот непосредственно в волокне, что в совокупности создает благоприятные условия для интенсификации процессов синтеза белка. При условии достаточного по времени и полноценного отдыха, отсутствия новых стрессовых нагрузок, адекватного снабжения волокна энергией и пластическими ресурсами (аминокислотами) интенсивные процессы восстановления приведут к накоплению в волокне белковых структур сверх уровня, который был до тренировки, то есть будет наблюдаться гипертрофия мышц.
Надо отметить, что последовательность протекания фаз общей неспецифической адаптационной реакции (синдрома стресса) такова, что обеспечивает поддержку описанных выше регенерационных процессов на системном уровне. Первая катаболическая фаза стресс реакции, сопровождается выбросом кортикостероидов, что приводит к мобилизации энергетических ресурсов организма и обеспечивает индукцию ферментов лизосом и фагоцитов, расщепляющих белок (кортикостероиды являются теми гормонами, которые активируют на ДНК клеток гены протеолитических ферментов), что способствует скорейшему очищению волокон от поврежденных структур. В последующей фазе стресс реакции синтез кортикостероидов сменяется синтезом анаболических гормонов, что обеспечивает на системном уровне компенсаторный анаболизм.
Как тут не вспомнить об основном законе философии — единстве и борьбе противоположностей. Анаболизм активируется катаболизмом — рост мышц есть следствие их предварительного разрушения.
Что такое микротравма
Открытым остается вопрос, что вызывает разрушение внутренней структуры волокна и является тем самым стрессом для мышцы? Прежде чем ответить на этот вопрос, рекомендую читателям вспомнить механизм сокращения мышц, описанный в I части.
Ученый и пауэрлифтер Фредерик Хетфилд, считающий роль микротравм в тренировочном процессе скорее отрицательной из-за необходимости длительного восстановления, полагает, что причиной микротравм является повреждение миофибрильных нитей во время негативных повторений. Вот как он описывает механизм этих повреждений: «Так как количество перекрестных мостиков, старающихся сократить мышцу недостаточно, они буквально «продираются» сквозь мостики соединений нити, стараясь вызвать концентрическое сокращение. Однако сцепиться, как следует им не удается, они срываются и повреждаются. Эти действия, очень напоминающие протаскивание щетины одной зубной щетки через другую, сопровождаются сильным трением, и мышечные нити разрушаются»
Не правда ли образно? Но, на мой взгляд, несколько сумбурно — так пишут когда хотят объяснить то, что до конца не понимают сами. К сожалению, мэтр ошибся дважды.
Во-первых, микротравмы возникают не только при негативных повторениях, но и при позитивном движении.
Во-вторых, использовать термин «трение» для описания взаимодействия молекул некорректно. Сила трения введена в физике для описания на макро-уровне поверхностного взаимодействия тел специально, дабы абстрагироваться от истинной природы «трения» — электромагнитного взаимодействия молекул поверхностного слоя.
Механизм повреждения миофибрильных нитей носит несколько иной характер, чем описывает Хетфилд, и мне удалось его аналитически смоделировать.
Для понимания механизма повреждения миофибрильных нитей следует обратиться к рассмотрению фаз движения миозинового мостика, которые вкратце уже описаны в первой части статьи. Сейчас остановимся подробнее на этом вопросе (рис. 8).
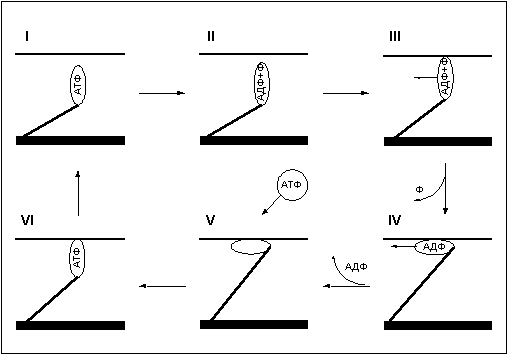
Рис. 8.
Итак, в первой фазе, еще до сцепления с актином, головка миозинового мостика несет в себе АТФ. Далее во второй фазе под действием фермента АТФаза АТФ гидролизуется, расщепляясь на АДФ и неорганический фосфат. Происходит это на не связанном с актином миозине, после этого миозиновая головка может соединятся с актином — третья фаза. Для совершения рабочего хода мостика используется энергия, освобождающаяся при диссоциации продуктов гидролиза АТФ. Основная доля энергии выделяется при высвобождении неорганического фосфата (переход из третей фазы в четвертую) и меньшая часть при высвобождении АДФ (переход из четвертой фазы в пятую). В пятой фазе -ригорное состояние мостика, мостик уже не генерирует силу, но по-прежнему находится в сцепленном состоянии, вывести его из этого состояния может только молекула АТФ. Поглощая АТФ, головка миозина переходит в шестую фазу, после чего отцепляется от актина, возвращаясь в исходное состояние (первая фаза).
Анализируя фазы движения миозинового мостика, я сразу обратил внимание на тот факт, что для отцепления мостика от актина требуется молекула АТФ. При скольжении нитей миозина вдоль актина под действием сил тянущих мостиков (позитивное движение) или под действием внешней силы (негативное движение) сцепленные мостики растягиваются и мешают движению, этим, как вы помните, объясняется различие в силе развиваемой волокном при удлинении и сокращении и сокращении с разной скоростью. Когда АТФ в мышце находится в достаточном количестве, мостики успевают вовремя отцепиться, но что будет, если, при снижении концентрации АТФ в мышце, молекула АТФ не успеет отцепить головку миозина до того, как растяжение мостика превысит предел его прочности? Естественно сцепленный мостик разорвется! (Рис. 9).
Вы, наверное, слышали о состоянии трупного окоченения мышц? Наступает оно потому, что в мертвом организме запасы АТФ не восполняются, и мостики миозина оказываются накрепко сцепленными с актином. Представьте, что произойдет с мышцей трупа, если ее насильно растянуть. Так вот нечто подобное происходит и с отдельными волокнами живой мышцы, в которых по причине интенсивной деятельности резко снижается уровень АТФ.
Надо понимать, что изображенное мной место разрыва мостика достаточно условно, я не могу точно сказать, где находится самое слабое звено в цепи, но то, что при движении, сопровождающемся недостатком АТФ, должно происходить нефизиологическое разрушение акто-миозинового комплекса — неоспоримо.
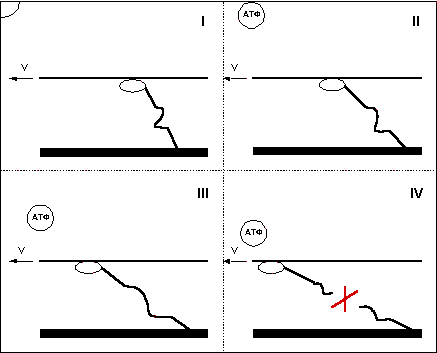
Рис. 9.
«Эврика!» — воскликнул я. Вот оно объяснение системы Ментцера! Работа до «отказа» вызывает истощение запасов АТФ и, соответственно, разрывы в акто-миозиновом комплексе, что является стрессом для мышцы и вызывает адаптационную реакцию. Сразу спешу предупредить читателей, радость моя была преждевременной. Как оказалось «отказ» наступает вовсе не из за исчерпания запасов АТФ, и уровень АТФ остается достаточно высоким даже в уставшей мышце. Решение задачи оказалось не таким простым, зато еще более красивым.
Сомневаться в правильности «отказной» теории, меня заставил и тот факт, что проявление боли в мышцах — вестника микротравм не имеет прямой связи с работой до «отказа». С одной стороны, на начальном этапе тренинга боль в мышцах возникает при любой работе, не зависимо от того был «отказ» или нет. С другой стороны регулярные тренировки приводят к тому, что даже работа сверх «отказа» (читинг, стриптиз и др.) не вызывает боли в мышцах — боль возвращается только после продолжительного перерыва в тренировках.
Эти противоречия заставили меня заняться детальным изучением процесса «отказа».
Что такое «отказ» мышцы?
«Отказ» понятие довольно условное, это не особое состояние мышцы, а ни что иное, как неспособность развить силу, необходимую для преодоления внешней нагрузки. Причиной отказа является снижение силы, генерируемой целой мышцей и отдельными волокнами. Сила отдельного волокна зависит от количества одновременно тянущих миозиновых мостиков. Биохимических причин снижения силы, развиваемой мостиком в генерирующих силу фазах (III, IV) не существует, во всяком случае, пока об этом ничего не известно, поэтому сила волокна может снизиться только по следующим причинам:
- Увеличение времени нахождения мостиков в ригорном — сцепленном не генерирующем силу состоянии (фаза V, VI);
- Увеличение времени нахождения мостиков в разомкнутом состоянии (фаза I, II).
- Увеличение времени нахождения мостиков в ригорном — сцепленном не генерирующем силу состоянии (фаза V, VI);
- Увеличение времени нахождения мостиков в разомкнутом состоянии (фаза I, II).
Вспоминая аналогию с гребцами, скажем, что скорость движения лодки и развиваемая при этом сила тем выше, чем быстрее весла вытаскиваются из воды после гребка и чем меньше гребцы сидят с поднятыми веслами без дела.
Время нахождения мостика в ригорном состоянии зависит от того, как быстро АТФ вступит в контакт с головкой миозина. Как известно частицы вещества постоянно находятся в хаотическом движении (рис. 10). Предположим что r — некий радиус, ограничивающий область пространства вокруг головки миозина, при попадании в пределы которого молекулы АТФ становится возможным реакция, приводящая к отцеплению головки от актина.
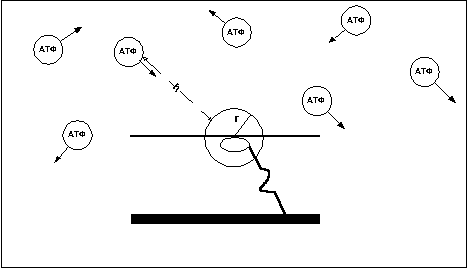
Рис. 10
В этом случае время нахождения мостика в ригорном состоянии равно частному от деления расстояния (h) от мостика до ближайшей молекулы АТФ, движущейся в направлении мостика, на cскорость движения молекулы (v). Среднее же время нахождения мостиков в ригорном состоянии равно средневзвешенному расстоянию между молекулами АТФ деленному на средневзвешенную скорость движения молекул.
Т=h/v
Естественно чем выше концентрация АТФ в мышце, тем меньше среднее расстояние между молекулами АТФ и головками миозина и тем меньше время нахождения мостиков в ригорном состоянии.
То есть сила волокна зависит от концентрации АТФ. При прочих равных условиях волокно с высокой концентрацией АТФ сильнее такого же волокна, но с меньшей концентрацией АТФ. Известно, что воспроизводство АТФ за счет креатинфосфата происходит почти мгновенно, поэтому можно сказать, что концентрация АТФ при работе мышцы зависит и от концентрации креатинфосфата. Таким образом:
Чем выше концентрация энергетических фосфатов (АТФ и креатинфосфата) в мышечном волокне, тем выше сила, развиваемая волокном.
После зависимости силы волокна от количества и поперечного сечения миофибрилл, влияние концентрации энергетических фосфатов на силу волокна является наиболее значимым. Именно по этой причине многие атлеты отмечают некоторую прибавку в силе, повышая уровень креатинфосфата в мышцах, принимая креатин, в качестве пищевой добавки.
Надо заметить, что зависимость силы волокна от концентрации энергетических фосфатов не является линейной. При высоких концентрациях, когда время ожидания контакта АТФ с головкой миозина станет меньше или сопоставимым со временем, необходимым для протекания химической реакции, приводящей к отделению головки миозина от актина, повышение концентрации АТФ и креатинфосфата будет мало влиять на скорость переключения мостиков и соответственно силу волокна. Зато низкие концентрации АТФ и креатинфосфата существенно снизят силу волокна, что может служить одной из причин отказа.
Рассмотрим второй фактор снижения силы сокращения — увеличение времени нахождения мостиков в разомкнутом состоянии (I, II фазы), что может происходить в результате снижения скорости гидролиза АТФ (снижения активности АТФазы — фермента ускоряющего реакцию гидролиза АТФ).
Расщепление АТФ и его воспроизводство за счет креатинфосфата вызывает накопление в мышце ортофосфорной кислоты, а гликолиз ведет к накоплению молочной кислоты. Активность АТФазы — фермента, благодаря которому происходит гидролиз АТФ, сильно зависит от кислотности среды. Как показывают эксперименты, максимум АТФазной активности достигается при среде близкой к нейтральной (рН=7), а при смещении среды мышцы в кислую сторону активность АТФазы падает, и при снижении рН среды до 5 АТФазная активность стремится к нулю (Богач П.Г. с соавторами, Поглазов Б.Ф). Таким образом, при накоплении кислых продуктов метаболизма АТФаза миозина постепенно снижает скорость гидролиза АТФ, и мостики теряют способность сцепляться с актином, при этом мышца снижает силу сокращения, несмотря на поступающий от мотонейрона сигнал.
Существует и еще как минимум две причины снижения силы мышц — при длительной работе накопление продуктов метаболизма тормозит процессы передачи сигнала от мотонейрона к волокну (Романовский Д.Ю.), а в центральной нервной системе развивается охранительное торможение.
То есть «отказ» мышцы является суперпозицией различных причин и доминирование одного либо другого фактора определяется интенсивностью работы мышцы.
Еще раз об энергетике
Резкая активизация мышечной деятельности из состояния покоя требует такого же резкого увеличения скорости производства энергии.
Для достижения максимальной мощности основных источников воспроизводства энергии (гликолиза в быстрых волокнах и окисления в медленных) требуется время.
Скорость воспроизводства АТФ за счет гликолиза достигает своего максимума только через 20 -30 секунд после начала интенсивной работы.
Для достижения максимальной скорости окислительного процесса требуется гораздо больше времени, связано это в основном с необходимостью оптимизацией процессов доставки кислорода. Скорость окисления становится максимальной лишь через 1-2 минуты работы мышц, этот эффект наверняка известен вам под названием «второе дыхание».
Между тем мышца развивает максимальную мощность с первых же долей секунд после поступления команды к сокращению, гликолиз, в совокупности с окислением, не в состоянии обеспечить необходимую скорость воспроизводства АТФ для поддержания этой мощности. Приведение в соответствие скоростей расхода и воспроизводства АТФ во время работы мышцы идет по двум направлениям. Во-первых, постепенная активизация гликолиза и окисления увеличивает количество АТФ, синтезируемого в единицу времени за счет этих источников. Во-вторых, накопление продуктов метаболизма, в результате деятельности гликолиза и окисления, снижает активность АТФазы миозина и соответственно скорость расхода АТФ. Благодаря этим двум процессам скорости расхода и воспроизводства АТФ выравниваются, и в дальнейшем движение продолжается с постепенно снижающейся мощностью, но в состоянии равновесия между количеством синтезируемого АТФ и потребностями мышцы в энергии. Отказ же мышцы наступает не из-за окончания запасов АТФ, а из-за снижения сократительной способности мышц в результате накопления кислых продуктов метаболизма.
До выравнивания скоростей расхода и воспроизводства энергии дефицит АТФ покрывается за счет имеющегося в мышце креатинфосфата. То есть креатинфосфат играет роль буфера энергии, сглаживающего несоответствия в скоростях воспроизводства и потребления АТФ при резко возрастающих нагрузках.
В обычной жизни мы редко используем собственные мышцы на пределе их энергетических возможностей, поэтому они вполне обходятся небольшим запасом креатинфосфата и ферментов обеспечивающих протекание реакций гликолиза и окисления, достаточным для повседневной жизни. По приходу в спортивный зал мышцы оказываются неготовыми к предстоящей работе. И если дать нагрузку, значительно превышающую привычную, то запас креатинфосфата в волокнах, первыми включившихся в работу, очень быстро заканчивается еще до того момента, когда процессы гликолиза в быстрых волокнах или окисления в медленных наберут обороты и обеспечат приемлемую скорость воспроизводства АТФ. Таким образом, из за интенсивного расхода и неадекватной скорости воспроизводства энергии, уровень АТФ в ряде волокон падает ниже критического. Так как движение продолжается под действием силы других волокон или внешней силы (при негативном движении), то в рассматриваемых нами волокнах происходит разрушение миофибрильных нитей.
Вот главный секрет тренировочного стресса: Микротравмы мышечного волокна возникают при исчерпании запасов креатинфосфата до того, как скорость воспроизводства АТФ за счет гликолиза и окисления станет равной скорости расхода АТФ.
Этим и объясняется тот факт, что тренировочный эффект воздействия на быстрые волокна достигается при интенсивной работе длительностью от 7 до 30 секунд. Если нагрузка позволяет поддерживать требуемую силу сокращения мышц дольше чем 30 секунд, то скорость расхода энергии в мышце, скорее всего, будет не достаточно велика для падения концентрации АТФ ниже критического уровня. Отказ мышцы в этом случае наступает в результате накопления кислых продуктов метаболизма, и является физиологически нормальным явлением, не оказывая на мышцу стрессового воздействия. Когда нагрузка велика, но может продлиться не дольше нескольких секунд (2-3 повторения), наблюдается другая картина. Скорость расхода энергии достаточно высока, но отказ, вызванный легким снижением силы волокон по причине накопления продуктов метаболизма и снижения концентрации АТФ (но не ниже критического уровня), происходит еще до исчерпания запасов креатинфосфата, и стрессовая ситуация не наступает.
Понятно, что при такого рода режиме работы мышц добиться микротравм в окислительных (медленных) волокнах невозможно. Скорость расхода АТФ в медленных волокнах значительно ниже, чем в быстрых, поэтому запасы креатинфосфата истощаются плавно. И, пожалуй, получение микротравм в медленных волокнах было бы практически невозможным, если бы для активации окислительных процессов требовалось столько же времени, как для активизации гликолиза. Но, как я упоминал ранее, максимум выработки АТФ за счет окисления наблюдается только через 1-2 минуты работы, поэтому есть шанс добиться микротравм в медленных волокнах если успеть получить дефицит АТФ в результате интенсивной работы в течении 1-2 минут.
Предложенная мною модель получения микротравм очень хорошо согласуется с еще одним физиологически важным явлением, известным каждому спортсмену, но до сих пор не получившему сколько ни будь приемлемого объяснения, — посттренировочная боль особенно сильна после первых занятий и практически полностью исчезает при регулярных тренировках, появляясь вновь только в случае длительного перерыва. Секрет этого явления очень прост — ответной реакцией на тренировку, помимо усиления синтеза белка, является накопление в мышце креатинфосфата и повышение концентрации и активности ферментов гликолиза и окисления. С каждой тренировкой относительное количество креатинфосфата в мышечном волокне увеличивается, растет и мощность гликолиза и окислительных реакций, в результате добиться исчерпания запасов креатинфосфата до выравнивания скоростей расхода и восстановления АТФ за счет основных источников энергии становится все труднее, а при высоком уровне тренированности практически невозможно.
И так, вот еще один важнейший вывод:
Накопление креатинфосфата и рост мощности гликолиза и окисления в результате тренировок, с одной стороны, повышает силу мышц и способствует росту их работоспособности, с другой стороны, препятствует созданию стрессовых ситуаций и снижает воздействие тренировки на мышцу, тем самым, замедляя дальнейшие адаптационные реакции.
Явление постепенного «привыкания» мышц к нагрузке известно методистам бодибилдинга достаточно давно под названием «тренировочное плато». До сих пор биохимические причины процессов, приводящих к снижению восприимчивости мышц к тренировке, были не известны, поэтому для преодоления плато чаще всего советовали сменить режим работы, заменить выполняемые упражнения, изменить тренировочный сплит, либо увеличить объем нагрузки, чтобы как то по новому воздействовать на мышцу и добиться от нее ответной реакции.
Между тем, для того, чтобы добиться снижения концентрации АТФ, по мере накопления в мышце большого количества креатинфосфата и роста ее энергетических возможностей необходимо постоянно повышать скорость расхода энергии, для чего в распоряжении атлета имеется не так много способов, од ин из них — повышение веса снаряда. Постоянное увеличение веса снаряда, в стремлении интенсифицировать нагрузку приводит к тому, что количество повторений в движении опускается ниже 4-х, что, как я показал выше, не может оказать на мышцу необходимого воздействия. Увеличение же объема работы за счет количества упражнений и подходов в этом случае малоэффективно. Если интенсивность расхода энергии не достаточно высока для исчерпания накопленного в мышце креатинфосфата до выравнивания скоростей расхода и синтеза АТФ, и стрессовая ситуация не наступает в первом же подходе, то последующие подходы, скорость расхода энергии в которых меньше чем в первом подходе, в виду остаточного накопления продуктов метаболизма, тем более не дадут нужного эффекта. Напоминаю, что уровень креатинфосфата в мышце восстанавливается в течение нескольких минут, а вот молочная кислота, снижающая мощность сокращения, полностью выводится из мышцы лишь в течение нескольких часов после тренировки. Таким образом, по мере приспособления мышц к тренировочным нагрузкам, тренировка из стрессового фактора превращается в обычную работу. В таком состоянии спортсмен может тренироваться почти каждый день, не наблюдая никакой перетренерованности, так как для восстановления мышц ему оказывается достаточным даже одного-двух дней отдыха, именно по тому, что такая тренировка не вызывает разрушения волокон. Но есть ли смысл в тренировках такого рода для бодибилдера? Да, при постепенном наращивании объема работы будет расти и объем саркоплазмы волокон, за счет накопления энергетических веществ, но такой рост не беспределен. Без увеличения количества и объема миофибрилл, а особенно количества клеточных ядер в волокнах (не будем исключать и возможность увеличения количества самих волокон), добиться значительной гипертрофии мышц невозможно.
Способность мышц сокращаться с требуемым усилием определяется, прежде всего, насыщенностью мышц энергией. И хотя основной причиной снижения силы сокращения мышц является вовсе не отсутствие АТФ, а снижение АТФазной активности миозина и нарушения в механизме передачи возбуждения с нерва вглубь волокна, причиной упомянутых нарушений являются метаболические факторы утомления (молочная кислота, ортофосфорная кислота, АДФ и др.), а их появление в мышце связано как раз с доступностью энергии. Недостаток АТФ, производимой окислительным путем, приводит к активизации гликолиза и появлению в мышце большого количества молочной кислоты (лактата), недостаток энергии, производимой путем гликолиза, приводит к истощению запасов креатнфосфата и, соответственно, увеличению в мышце концентрации ортофосфата.
По Н.И. Волкову при рассмотрении факторов работоспособности мышц, в зависимости от основного механизма энергообеспечения, следует различать аэробную (окисление) и анаэробную работоспособность, а анаэробная работоспособность, в свою очередь, делится на лактатную (гликолиз) и алактатную (креатинфосфат). В качестве главных критериев оценки механизмов энергообеспечения мышечной деятельности принято выделять максимальную мощность, время удержания максимальной мощности, и общую емкость механизма. Максимальная мощность — это наибольшая скорость образования АТФ в данном метаболическом процессе. От мощности механизма энергообеспечения зависит возможная сила сокращения мышц в данном режиме работы. Под емкостью понимается общее количество энергии, которое можно получить за счет данного механизма ресинтеза АТФ.
Алактатная работоспособность мышц
Максимальная алактатная мощность, с одной стороны, зависит от концентрации и активности фермента креатинкиназа (переносящего фосфатную группу с креатинфосфата на АДФ) и собственно креатинфосфата, с другой стороны мощность данной реакции зависит от потребности мышц в энергии, соответственно, определяется максимальной скоростью расхода АТФ развиваемой мышцами. Максимальная длительность удержания алактатной мощности составляет 6-12 секунд. Алактатная емкость зависит от запасов креатинфосфата в мышце. О методах тренировки алактатной мощности и емкости я уже рассказывал ранее, рассматривая методы развития максимальной силы, и сейчас не буду подробно останавливаться на этом вопросе.
Лактатная работоспособность мышц
Максимальная лактатная мощность определяется главным образом конценрацией и активностью ключевых ферментов гликолиза. Время удержания максимальной мощности данного метаболического процесса составляет 30-60 секунд, и определяется, с одной стороны, устойчивостью ферментов гликолиза к понижению рН среды (повышение кислотности среды ингибирует активность гликолитических ферментов, что подавляет энергопроизводство), и устойчивостью кислотно-щелочного равновесия внутренней среды мышц, в условиях усиленной выработки лактата. С другой стороны, время удержания максимальной гликолитической мощности лимитируется факторами утомления мышцы, снижающими интенсивность сокращения.
Из вышесказанного следует, что для запуска адаптационных процессов, направленных на увеличение максимальной гликолитической мощности, длительность нагрузки должна соответствовать времени удержания максимальной мощности данного метаболического процесса, что составляет 30-60 секунд. Отдых между подходами должен быть достаточно длительным, для обеспечения вывода продуктов метаболизма из мышцы и развития высокой мощности гликолиза в следующем подходе. Устойчивость рН среды мышечных волокон к выбросу молочной кислоты и устойчивость ключевых ферментов к снижению рН вырабатывается в ходе тренировок, сопровождающихся максимальным накоплением лактата в мышцах. Это могут быть нагрузки высокой интенсивности, длительностью 1-1.5 минуты до наступления отказа мышц, вызванного сильным закислением, либо более короткие нагрузки, длительностью 20-40 секунд, со столь же коротким интервалом отдыха, приводящие к кумулятивному накоплению лактата в мышцах.
Гликолитическая емкость определяется главным образом запасами гликогена в мышцах, гликоген печени для процессов гликолиза не обладает достаточной мобильностью. О методах накопления мышечного гликогена, как и гликогена печени, я уже рассказывал при рассмотрении факторов общей работоспособности организма.
Аэробная работоспособность мышц
Максимальная аэробная мощность зависит главным образом от плотности митохондрий в мышечных волокнах, концентрации и активности окислительных ферментов, скорости поступления кислорода вглубь волокна. Объем кислорода доступного для окислительных реакций лимитируется, как факторами общей работоспособности организма, которые я уже ранее рассматривал, так и рядом локальных внутримышечных факторов, среди которых можно выделить капиляризацию мышц, концентрацию миоглобина, диаметр мышечного волокна (чем меньше диаметр волокна, тем лучше оно снабжается кислородом и тем выше его относительная аэробная мощность). Скорость производства АТФ за счет окисления достигает максимальных значений на 2-3-й минуте работы, что связано с необходимостью развертывания множества процессов, обеспечивающих доставку кислорода к митохондриям. Время удержания максимальной аэробной мощности составляет примерно 6 минут, в дальнейшем аэробная мощность снижается по причине усталости всех активно работающих систем организма. Соответственно, для повышения аэробной мощности мышц тренировочная нагрузка должна длиться не менее 2 минут (для выхода скорости энергопроизводства на максимум). Не имеет смысла и затягивать нагрузку дольше чем на 6 минут, при тренировке именно мощности, так как далее идет ее (мощности) снижение. Эффективным оказывается многократное повторение таких нагрузок.
В заключение хочу привести сводную таблицу тренировочного воздействия на работоспособность мышц в различных режимах работы, почерпнутую мной из диссертации М. Хосни, посвященной изучению биохимических основ интервальной тренировки. Для развития соответствующих качеств Хосни рекомендует следующие методические приемы:
| Направление воздействия тренировки | Интенсивность | Длительность нагрузки | Отдых между подходами | Количество подходов |
| Алактатная анаэробная мощность | Максимальная | 7-10 с. | 2-5 мин. | 5-6 |
| Алактатная анаэробная емкость | Максимальная | 7-10 с. | 0.3-1.5 мин. | 10-12 |
| Лактатная анаэробная мощность | Высокая | 20-30 с. | 6-10 мин. | 3-4 |
| Лактатная анаэробная емкость | Высокая | 40-90 с. | 5-6 мин. | 10-15 |
| Аэробная мощность | На максимуме потребления кислорода | 0.5-2.5 мин. | 0.5-3 мин. | 10-15 |
| Аэробная емкость | На максимуме потребления кислорода | 1-6 мин. | 1-6 мин. | Больше 10 |
Ещё раз огромное спасибо Вадиму! Те кто дочитал до этого момента могут смело собой гордиться — теперь они знают о теории сокращения мышц больше чем «инструктор» в фитнес зале. Тех читатели чьей главной целью является построение дополнительных мышечных объемов я приглашаю прочесть полный текст книги Вадима Протасенко, к счастью она доступна в свободном доступе. Всех остальных я поздравляю после прочтения этого поста вы готовы к последующим публикациям, в которых мы с вами будем детально будем рассматривать и классифицировать современные модные тенденции построения красивого но и мощного и всесторонне развитого тела а также стратегии снижения веса, то есть функциональный тренинг и похудение.
Комментариев нет:
Отправить комментарий