ФИО первого автора:
Елдаров Адам Вахитович
Место его работы или учебы:
студент Московского государственного университета экономики, статистики и информатики (МЭСИ), РФ, г. Москва
Ф.И.О. научного руководителя:
Буянова Татьяна Владимировна
Место работы руководителя, степень, звание, ВУЗ, город, страна:
преподаватель Московского государственного университета экономики, статистики и информатики (МЭСИ), РФ, г. Москва
В современной фитнес-индустрии существует огромное множество тренировочных методик, предназначенных для огромного спектра задач. Но мы рассмотрим в статье все, что связанно с развитием мускулатуры и мышечной гипертрофии, в частности.
К сожалению, авторы большинства методологий не имеют четкого представления о причинах роста мышц. Они либо игнорируются, либо искажаются. Поистине, фитнес-индустрия собирает вокруг себя огромное количество мифов, стереотипов и заблуждений. Порой, задача получить качественную информацию становится весьма и весьма непростой. В этой статье мы попытаемся разобраться с этим вопросом.
Мышечная гипертрофия — увеличение размеров мышечного волокна.
Как известно, все мышцы состоят большого количества мышечных волокон, которые крепятся к одному сухожилию, образуя при это, так называемые пучки.
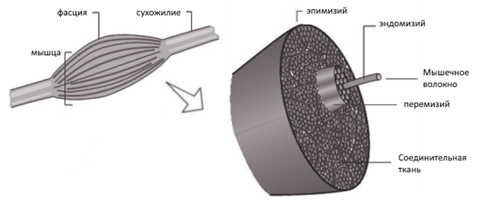
Рисунок 1. Анатомия мышцы
Мышечное волокно (мышечная клетка, миоцит) состоит из миофибрилл, саркоплазматического пространства, митохондрий, ядер и т. д. Из себя представляет вытянутую клетку, которая способна сокращаться, благодаря сокращению нитевидных миофибрилл, состоящих из белков двух типов: актина и миозина. В саркоплазме же находятся энергозапасы клетки: Креатин фосфат, гликоген, ферментативные белки, соли, вода и т. д. [1, c. 14].
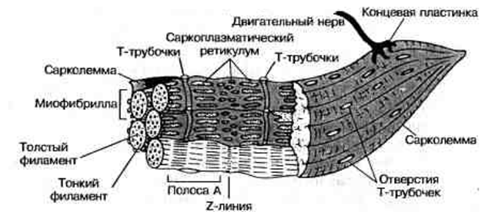
Рисунок 2. Анатомия мышечного волокна
ВИДЫ МЫШЕЧНЫХ ВОЛОКОН.
Мышечные волокна бывают нескольких видов: медленные (тип I) и быстрые (тип II) [2].
Существует мнение, что медленные волокна работают при медленных движениях, но это не верно, так как классификация на медленные и быстрые основывается на активности АТФазы (фермент необходимый для мышечного сокращения): чем выше активность, тем мощнее сокращение. У медленных волокон скорость АТФазы гораздо ниже. У каждого из этих видов есть подтипы. Так же волокна различаются по типу энергообеспечения: окислительные и гликолитические. Окислительные — означает, что работает за счет окисления жирных кислот и глюкозы и для их работы необходим кислород, а гликолитические работают на анаэробном (без доступа кислорода) гликолизе. Окислительные волокна более выносливы и наименее сильные, а гликолитические имеют крайне малую длительность работы (около минуты), но обладают наибольшей мощностью и силой сокращения [5, c. 32].
В Таблице 1 сверху вниз перечислены типы волокон в зависимости от активности их АТФазы. Волокна типа I наименее мощные и сильные, а IIB имеют самый большой силовой потенциал и сокращение их наиболее мощное.
Таблица 1.
Классификация мышечных волокон
Тип волокон
|
Тип энергообеспечения
|
I
|
SO
|
IC
| |
IIC
|
FOG
|
IIAC
| |
IIA
| |
IIAB
| |
IIB
|
FG
|
*SO (slow-oxidative) — медленные окислительные, FOG (fast-oxidative/glycolytic) — быстрые окислительно-гликолитические, FG (fast-glycolytic) — быстрые гликолитические
Так же, в таком порядке происходит иннервация, то есть включение в работу во время сокращения мышцы. Чем выше сигнал подается к мышце, тем больше волокон получают нагрузку. Самый низкий порог иннервации у медленных волокон I (работают, когда мы ходим), наиболее высокий у быстрых (спринт или работа с большими отягощениями). Мощность сигнала зависит от нескольких показателей: интенсивность (процент отягощения от разового максимума), скорость сокращения и доступ кислорода. Чем выше интенсивность, тем больше волокон включается в работу [5, c. 32]. Опыты Роджера Эноки показали, что при отягощении с 75 % от 1ПМ (повторный максимум) и выше, работают все единицы, а при быстром темпе, хватает и 40 % от 1ПМ [9]. Так же важен доступ кислорода. Если доступ его ограничен (достигается за счет перекрытия кровотока во время выполнения упражнения), то из работы выключаются окислительные волокна (так как нужен кислород для работы) и включаются более высоко пороговые гликолитические, которые могут работать в условиях гипоксии. Перекрытие кровотока достигается за счет отягощения (чем выше оно, тем сильнее гипоксия), укорачивания амплитуды движения и специальных приспособлений (жгуты). Опыты показывают, что при весе 20 % 1ПМ и перетягивании целевой мышцы жгутом, работают все двигательные единицы [3; 4].
Разобравшись с теоретической основной, поговорим про развитие мускулатуры.
МЫШЕНЧАЯ ГИПЕРТРОФИЯ.
Мышечная гипертрофия может быть определена, как увеличение размера волокна за счет накопления сократительных или не сократительных белков, который происходит за счет увеличения синтеза белка после силовых тренировок, снижения распада белка или сочетания этих двух факторов, на которые можно влиять физическими тренировками [5, c. 33].
Существует три тренировочных стимула, вызывающих мышечную гипертрофию: Механический натяг, микротравмы и метаболический стресс [6, c. 5].
Рассмотрим каждый из стимулов немного подробнее:
МЕХАНИЧЕСКИЙ НАТЯГ (MECHANICAL TENSION).
Является следствием нарушения целостности волокна во время генерации силы или его растяжения. В результате чего провоцируется ответ на клеточном и молекулярном уровне: модуляция гормонов (Инсулиноподобный фактор роста-1, механический фактор роста), белков регуляторов, увеличение транскрипции иРНК. Основным регулятором данного процесса является комплекс AKTmTOR, как мы можем видеть на рисунке 3. Стоит учесть, что идет стимулирование развития сократительного аппарата во всех типах волокон — миофибрилл [6, c. 6].
МИКРОТРАВМЫ.
Физические тренировки могут привести к локальному повреждению мышц, которые при определенных условиях могут создать гипертрофический ответ. Ущерб может быть специфичный для всего несколько макромолекул ткани, или привести к разрывам в сарколемме, базальной мембране и поддержку соединительной ткани и стимулирует травмы сократительных элементов и цито скелета. Считается, что это приводит к высвобождению различных ростовых факторов, которые регулируют пролиферацию и дифференцировку клеток спутников (Миосателлиты — стволовые клетки миоцитов). Происходит увеличение, как сократительных белков, так и энзимных [6, c. 6].
Микротравмы характерны для физических нагрузок с искусственно завышенными весами, использованию негативных повторений, чрезмерному объему механического натяга, растяжкам. Относится ко всем типам волокон.
МЕТАБОЛИЧЕСКИЙ СТРЕСС.
Метаболический стресс проявляется в результате физической деятельности, которая полагается на анаэробный гликолиз для производства АТФ, в результате чего последующее накопление метаболитов, таких как лактат, ионы водорода, неорганический фосфата, свободный креатин и др. Мышечная ишемия, как было показано, необходима, чтобы произвести значительный метаболический стресс, и производит увеличение гипертрофического эффекта.
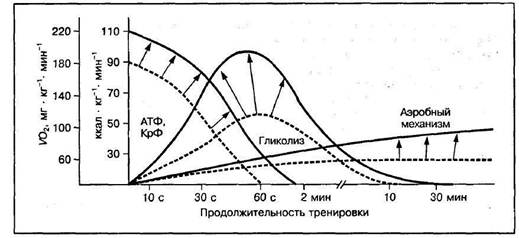
Рисунок 4. Адаптационные изменения мощности (VO2) и емкости отдельных механизмов энергообеспечения мышечной работы в процессе специфической тренировки
Метаболический стресс приводит к активации ростовых факторов, гормонов, белков активаторов, чувствительных к времени под нагрузкой, ферментов. Стимулирует рост сократительной части клетки, энзимов и запасов энергоресурсов клетки, а также увеличению количества воды внутри клетки [6, c. 6].
Стоит отметить, что данный стимул относится в большей мере к быстрым мышечным волокнам, а не к медленным, так как последние не обладают ферментами для анаэробного гликолиза.
Хотелось бы заострить немного внимания на таком ферменте, как АМФК. АМФК — клеточная протеинкиназа, контролирующая энергетический баланс клетки. Активируется при значительном потреблении энергии клетки. К состояниям, увеличивающим дефицит энергии в клетке и, соответственно, повышающим уровень АМПК, относятся физическая нагрузка, голод, гипоксия, ишемия, окислительный стресс и тепловой шок [7]. АМФК по сути триггер, который следит, чтобы клетка не умерла от голода. Важной её особенностью является то, что она блокирует синтез белка в клетке до тех пор, пока не восстановит ее энергетику [8]. Это важно знать, и мы вернемся к этому чуть позже.
МЕХАНИЗМ ГИПЕРТРОФИИ.
Рост сократительных элементов клетки проходит в несколько этапов [6, c. 3; 5, c. 102].
· Сначала мы задаем стимул с помощью физической нагрузки. Стимулы все ведут себя по-разному, но в конечном итоге происходит следующее.
· Под воздействием стимула происходит экспрессия иРНК (информационная РНК) внутри клетки, которая представляет из себя матрицу белков, некий чертеж или инструкцию.
· Далее иРНК направляется в рибосомы клетки, где они, руководствуясь этой матрицей синтезируют белки, собирая их из аминокислот, которые хранятся в клетке. В конечном итоге мы получаем увеличение мышечной массы.
Дело в том, что иРНК имеет период своей жизни, а рибосомы не могут быть бесконечно активными и имеют порог своей активность, как по времени, так и по абсолютным показателям. И, как следствие, синтез мышечного белка бывает повышен всего пару суток, а далее возвращается к исходным показателям, как видно на рисунке 5 [5, c. 103; 10].
Так же, мышечная гипертрофия будет наблюдаться в случае воздействия метаболического стимула, посредством увеличения гликогена, ферментных белков и задержки воды, тем самым в клетке больше будет «топлива», и они будут выглядеть более наполненными и выпуклыми.
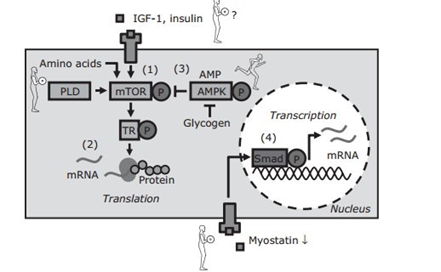
Рисунок 3. Схематическое представление стимулирования синтеза белка
Стоит обратить внимание на то, что показатели синтеза белка будут меняться в зависимости от тренировки, ее объема, интенсивности и прочих параметров, которые влияют на качество стимулирующего отклика. Так же не стоит забывать, что синтез белка является весьма энергозатратной процедурой и может падать, если в клетке будет недостаточно энергии, в следствии блокирующего действия АМФК [5, c. 104; 8].
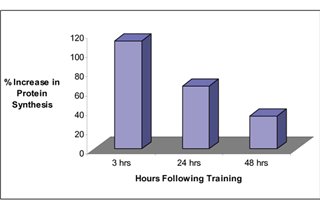
Рисунок 5. Возрастание синтеза белка в процентах после силовой тренировки
МЕДЛЕННЫЕ МЫШЕЧНЫЕ ВОЛОКНА (ММВ).
Медленные волокна воистину являются предметом для большого кол-ва мифов. Один из мифов — медленные движения, мы рассмотрели ранее. Второй же — это длительная нагрузка в условиях гипоксии (отсутствия кислорода) для их гипертрофии, особые стимулы и отсутствие эффекта при силовых тренировках с весами 60 % ПМ и выше. Данные утверждения являются ложными, потому что, как мы сказали ранее, ммв не имеют ферментов для анаэробного гликолиза и работают в условиях гипоксии только за счет запасов креатин фосфата, а это не более 10 секунд мощной и активной работы [2]. На рисунке 6 видно, что мощность работы в таком случае крайне мала по сравнению с полноценной анаэробной работой, а в условиях запроса на работу подключаются более высоко-пороговые двигательные единицы, которые способны работать в условиях гипоксии, то есть быстрые мышечные волокна. Это и демонстрируют исследования, как говорилось ранее. Даже с крайне низким весом (20 % ПМ) идет работа всех волокон, а гипертрофируются только быстрые [11, c. 69].
В тоже время, исследования показывают [12], что ммв прекрасно гипертрофируются при силовых тренировках с 60 % ПМ и выше, как это видно в таблице 2. Лучшие показатели роста волокон всех видов показала группа, использовавшая низко-повторный и средней-повторный тренинг.
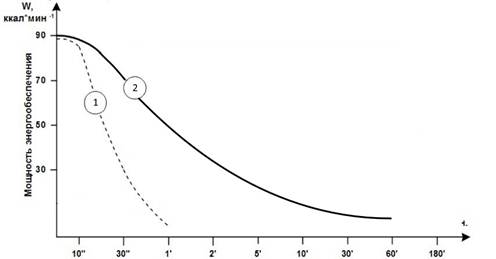
Рисунок 6. Мощность работы волокна при наличии ферментов для анаэробного гликолиза «2» и при их отсутствии «1»
Таблица 2.
Изменение площади (мкм2) поперечного сечения тренируемой мышцы в результате различных нагрузок
Тип тренинга
|
Тип волокон
| ||
I
|
IIA
|
IIB
| |
3—5 повторений по 4 подхода с весом 90 % ПМ. 3 минуты отдыха
| |||
До
|
4869
|
5615
|
4926
|
После
|
5475
|
6903
|
6171
|
9—11 повторов по 3 подхода с весом 75 % ПМ. 2 минуты отдыха
| |||
До
|
4155
|
5238
|
4556
|
После
|
4701
|
6090
|
5798
|
20—28 повторений по два подхода с отдыхом в 1 минуту и весом 30 % ПМ
| |||
До
|
3894
|
5217
|
4564
|
После
|
4297
|
5633
|
5181
|
ВЫВОДЫ.
В этой статье мы попытались разобраться, как растут мышцы. Стоит отметить, что на самом деле все куда сложнее и не так просто, но данной информации должно быть достаточно, чтобы получить теоретический фундамент для построения удачного тренировочного комплекса, целью которого будет развитие мускулатуры, но это тема для отдельной статьи.
Список литературы:
1. Мохан Р. Биохимия мышечной деятельности и физической тренировки / Глессон М., Гринхафф П.Л.; перевел с английского — Смульский В.Л. — Киев: Олимпийская литература, 2001 — 148 с.
2. Scott W., Stevens J., Binder–Macleod S.A. Human Skeletal Muscle Fiber Type Classifications // Physical Therapy November 2001 vol. 81 no. 11 1810—1816.
3. Wernbom M., Järrebring R., Andreasson M.A., Augustsson J. Acute effects of blood flow restriction on muscle activity and endurance during fatiguing dynamic knee extensions at low load. // J Strength Cond Res. 2009 Nov; 23 (8): 2389—95.
4. Fry C.S., E.L. Glynn, M.J. Drummond, K.L. Timmerman, S. Fujita, T. Abe, S. Dhanani, E. Volpi, B.B. Rasmussen. Blood flow restriction exercise stimulates mTORC1 signaling and muscle protein synthesis in older men. Journal of Applied Physiology, 2010; 108 (5): 1199 DOI: 10.1152/japplphysiol.01266.2009.
5. Cardinale M. Strength and Conditioning: Biological Principles and Practical Applications / Cardinale M., Rob Newton R., Kazunori Nosaka.; Oxford: John Wiley & Sons, Ltd, 2011 — 483 с.
6. Schoenfeld B.J. The mechanisms of muscle hypertrophy and their application to resistance training. J Strength Cond Res 24 (10): 2857—2872; 2010.
7. Witczak C.A., Sharoff C.G., Goodyear L.J. (September 2008). “AMP-activated protein kinase in skeletal muscle: From structure and localization to its role as a master regulator of cellular metabolism”. Cell. Mol. Life ScD.
8. The mechanism, by which AMPK inhibits protein synthesis potentially depends on down- and up-regulation of mTOR and eEF2 kinase signalling to 4E-BPs and eEF2, respectively (Horman et al. 2002; Inoki et al. 2003; Chan et al. 2004).
9. Enoka R.M. Modulation of motor unit activity in biceps brachii in response to neuromuscular electrical stimulation applied to the contralateral arm // Conference: European College of Sport Science, Amsterdam, 2014.
10. Phillips SM1, Tipton KD, Aarsland A, Wolf SE, Wolfe RR. Mixed muscle protein synthesis and breakdown after resistance exercise in humans. // Am J Physiol. 1997 Jul; 273 (1 Pt 1):E99 — 107.
11. T. Yasuda, T. Abe, Y. Sato, T. Midorikawa, C.F. Kearns K. Inoue, T. Ryushi, N. Ishii Muscle fiber cross-sectional area is increased after two weeks of twice daily KAATSU-resistance training // Int. J. KAATSU Training Res. 2005; 1: 65—70.
12. Gerson E.R. Muscular adaptations in response to three different resistance-training regimens: specificity of repetition maximum training zones // Accepted: 14 June 2002 / Published online: 15 August 2002.
1. Мохан Р. Биохимия мышечной деятельности и физической тренировки / Глессон М., Гринхафф П.Л.; перевел с английского — Смульский В.Л. — Киев: Олимпийская литература, 2001 — 148 с.
2. Scott W., Stevens J., Binder–Macleod S.A. Human Skeletal Muscle Fiber Type Classifications // Physical Therapy November 2001 vol. 81 no. 11 1810—1816.
3. Wernbom M., Järrebring R., Andreasson M.A., Augustsson J. Acute effects of blood flow restriction on muscle activity and endurance during fatiguing dynamic knee extensions at low load. // J Strength Cond Res. 2009 Nov; 23 (8): 2389—95.
4. Fry C.S., E.L. Glynn, M.J. Drummond, K.L. Timmerman, S. Fujita, T. Abe, S. Dhanani, E. Volpi, B.B. Rasmussen. Blood flow restriction exercise stimulates mTORC1 signaling and muscle protein synthesis in older men. Journal of Applied Physiology, 2010; 108 (5): 1199 DOI: 10.1152/japplphysiol.01266.2009.
5. Cardinale M. Strength and Conditioning: Biological Principles and Practical Applications / Cardinale M., Rob Newton R., Kazunori Nosaka.; Oxford: John Wiley & Sons, Ltd, 2011 — 483 с.
6. Schoenfeld B.J. The mechanisms of muscle hypertrophy and their application to resistance training. J Strength Cond Res 24 (10): 2857—2872; 2010.
7. Witczak C.A., Sharoff C.G., Goodyear L.J. (September 2008). “AMP-activated protein kinase in skeletal muscle: From structure and localization to its role as a master regulator of cellular metabolism”. Cell. Mol. Life ScD.
8. The mechanism, by which AMPK inhibits protein synthesis potentially depends on down- and up-regulation of mTOR and eEF2 kinase signalling to 4E-BPs and eEF2, respectively (Horman et al. 2002; Inoki et al. 2003; Chan et al. 2004).
9. Enoka R.M. Modulation of motor unit activity in biceps brachii in response to neuromuscular electrical stimulation applied to the contralateral arm // Conference: European College of Sport Science, Amsterdam, 2014.
10. Phillips SM1, Tipton KD, Aarsland A, Wolf SE, Wolfe RR. Mixed muscle protein synthesis and breakdown after resistance exercise in humans. // Am J Physiol. 1997 Jul; 273 (1 Pt 1):E99 — 107.
11. T. Yasuda, T. Abe, Y. Sato, T. Midorikawa, C.F. Kearns K. Inoue, T. Ryushi, N. Ishii Muscle fiber cross-sectional area is increased after two weeks of twice daily KAATSU-resistance training // Int. J. KAATSU Training Res. 2005; 1: 65—70.
12. Gerson E.R. Muscular adaptations in response to three different resistance-training regimens: specificity of repetition maximum training zones // Accepted: 14 June 2002 / Published online: 15 August 2002.
Комментариев нет:
Отправить комментарий